I. Spesifikt utfører Alu-sekvenser mange artsspesifikke, lavere-nivå formaterings-funksjoner, slik som :
i) forsyne alternative startsteder for aktivator-moduler i genuttrykk (kan sammenlignes med sektordeling på en harddisk; Faulkner et al (2009), Faulkner og Carnici (2009)
ii) undertrykke eller stilne RNA-avskrivning (Trujillo et al, 2006)
iii) dynamisk partisjonering av en gen-fil fra en annen på kromosomet (Lunyak et al, 2007)
iv) Frembringe DNA-knutepunkt for signal overføring, fordeling eller tilknytningssteder for hormon-reseptorer (Jacobsen et al, 2009; Laperriere et al, 2004)
v) Kode RNA som modulerer avskrifter (Allen et al, 2004; Espinoza et al, 2004; Walters et al, 2009)
vi) Kode eller regulere micro-RNAs (Gu et al, 2009; Lehnert et al, 2009)
Bilde 1. Antisense RNA hindrer oversettelse |
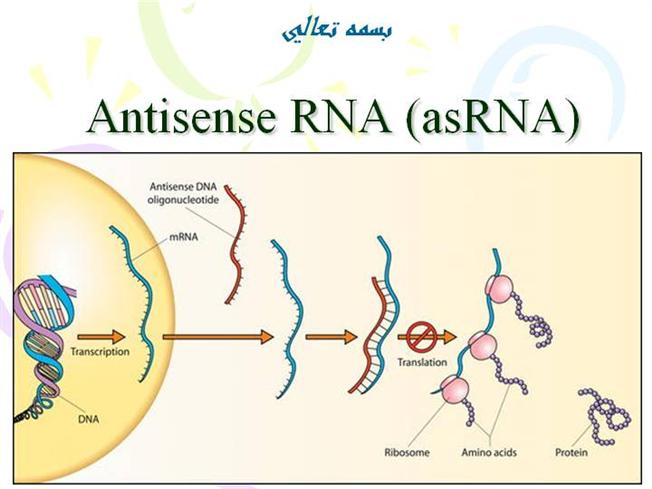 |
I tillegg til disse lavnivå formaterings-funksjonene i genomet, så utfører SINE-inkl. Alu, også spesifikke høyer-nivå genom-formaterings funkskjoner, slik som: |
Bilde 1. Eks. på RNA som blokkerer protein-oversettings maskineriet |
1) Modulering av kromatin for GC-rike signal-omformings og husholdnings (housekeeping) gener. (Groover et al. 2003; Oei et al, 2004)
2) 'Strek-koding' av spesielle segmenter for kromatin-looper mellom aktivator og forsterker elementer (Ford og Thanos, 2010)
3) Forstørre rekombinasjon i sekvenser der Alu-er inntreffer (Witherspoon et al, 2009)
4) Assistere i formasjon av tre-dimensjonale kromosom-seksjoner i genomet (Kaplan et al, 1993; Pai og Engelke, 2010)
Bilde 2. Kromatin-Proteomics |
 |
I tillegg spesifiserer Alu-sekvenser mange arts-spesifikke RNA-koder. Spesielt forsyner de: |
|
i) Signaler for arts-spesifikk spleising. De genererer flerfoldige mRNA fra samme type avskrift (Gal-Mark et al, 2008; Lei og Vorechovsky, 2005; Lev-Maor et al, 2008)
ii) alternative åpen-lesnings rammer (eksoner. Lev-Maor et al, 2007; Lin et al, 2008; Schwarz et al, 2009)
iii) Spesifikasjon av unndragelse av RNA i kjernen for å stilne gen-uttrykk (Chen et al, 2008; Walters et al, 2009)
iv) Regulerer RNA-polymerings maskineri i løpet av avskrift (Mariner et al, 2008; Yakovchuck et al, 2009; Walters et al, 2009)
v)Steder der Adenine-til-Inosine RNA-editering kan foregå.
Det er essensielt for menneskelig utvikling og arts-spesifikk hjerne-utvikling (Walters et al, 2009)
Bilde 3. RNA-editering kan påvirke mye |
 |
II. I motsetning til hva det hevdes fra darwinistisk hold, blir ikke SINE (inkl. Alus) fordelt tilfeldig, men synliggjør i stedet et 'strek-kode' mønster gjennom kromosomet. Fordeling av Alu-sekvenser og andre SINE's reflekterer en klar funksjonell logikk Chen og Manuelidis, 1989; Gibbs et al, 2004; Korenberg og Rykowski, 1988)
F.eks. foretrekkes Alu-sekvenser i og omkring protein-kodende gener, i det som passer i deres rolle i å regulere gen-uttrykk. (Tsirigos og Rigoutsos, 2009). De inntreffer hovedsakelig i aktivator regioner, start-stedene for RNA-produksjon, og i introner -segmentene som bryter opp protein-kodings strekkene. Utenfor disse områdene faller antall Alu's kraftig. Se mer på: Ayala and Falk Miss the Signs in the Genome.
Videre vet vi at Alu-sekvenser blir styrt -eller spleiset inn i visse foretrukne 'hotpots', i genomet av integerings-maskineriet tilhørende cellens informasjons-prosesserings system. ( Levy et al, 2010). Denne styrte fordelingen av Alu-sekvenser forsterker semantisk og syntaktisk organisering av menneskelig DNA. Det synes i det hele å ha lite å gjøre med tilfeldige innsettinger.
Bilde 4. Alus inntreffer ikke tilfeldig |
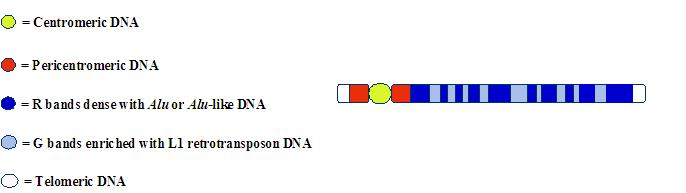 |